Mikroglej
 | Ten artykuł od 2014-01 zawiera treści, przy których brakuje odnośników do źródeł. Należy dodać przypisy do treści niemających odnośników do źródeł. Dodanie listy źródeł bibliograficznych jest problematyczne, ponieważ nie wiadomo, które treści one uźródławiają. Sprawdź w źródłach: Encyklopedia PWN • Google Books • Google Scholar • Federacja Bibliotek Cyfrowych • BazHum • BazTech • RCIN • Internet Archive (texts / inlibrary) Po wyeliminowaniu niedoskonałości należy usunąć szablon {{Dopracować}} z tego artykułu. |
Mikroglej (ang. microglia, third element) – komórki nieneuronalne centralnego układu nerwowego, tkankowo specyficzne, rezydentne makrofagi kontrolujące homeostazę i biorące udział w odpowiedzi immunologicznej.
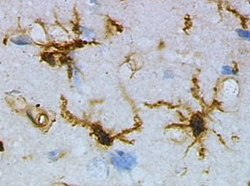
Historia
Pierwszymi poważnymi badaniami nad opisem gleju zajął się Santiago Ramon y Cajal (1852–1934) który scharakteryzował trzy podstawowe rodzaje komórek glejowych. Doniósł on o obecności komórek o kształcie gwieździstym – nieświadom tego że są to astrocyty, komórek włóknistych z licznymi wypustkami i komórek protoplazmatycznych występujących licznie w istocie szarej. Jak się okazało później jest to mikroglej. Niezależnie odkryty przez Nissla i Robertsona, dokładniejszy opis zyskał jednak dzięki uczniowi Cajala, Pío del Río-Hortedze (1882–1945). W wyniku barwienia solami srebra Hortega wyróżnił specyficzny rodzaj komórek zawierających jądro o wyraźnie wydłużonym kształcie. Scharakteryzowane przez niego komórki mikrogleju zyskały od tej pory miana komórek Hortegi. Następny „krok milowy” w badaniach nad mikroglejem należał znów do Hortegi, który opisywał rolę odkrytego mikrogleju w patologii centralnego układu nerwowego. W późniejszych latach, w wyniku zidentyfikowania komórkowo specyficznych markerów narastały kontrowersje wokół rozwoju embrionalnego jak i pochodzenia mikrogleju. Te i inne wątpliwości rozwiały zaawansowane badania w latach 80. z użyciem markerów lektynowych oraz rozwiniętych technik immunocytochemicznych.
Pochodzenie
Na temat pochodzenia mikrogleju prowadzono już wiele debat, na podstawie których nawet obecnie trudno jednoznacznie określić rodowód tych komórek[1]. Zdania są podzielone i część badaczy stoi na stanowisku, że źródłem mikrogleju są komórki linii monocytarnej, a więc prekursorów dopatrują się w mezodermie. Dowodem popierającym mezodermalną tezę są badania prowadzone przez Ling (Ling i in. 1980) polegające na znakowaniu monocytów z krwi obwodowej noworodków koloidalnym węglem i późniejszym lokalizowaniu ich w tkance nerwowej mózgu. Niepodważalnym dowodem jest również fakt, że komórki mikrogleju posiadają na swojej powierzchni markery F4/80, Mac-1, ED1, lektyny (GSA I-B4) typowe dla monocytów i makrofagów oraz receptory na Fc,CR3 (Perry i in. 1985). Faktem potwierdzającym założenie mezodermalnej teorii jest również obecność w cytoplazmie mikrogleju elementów lysozymu, oraz takich enzymów jak niespecyficzna esteraza czy peroksydaza.
Zgodnie z mezodermalną teorią pochodzenia komórek mikrogleju monocyty infiltrują parenchymę mózgu we wczesnych okresach życia embrionalnego jako mikroglej amebowaty. Zaznaczyć trzeba że napływ mikrogleju do mózgu jest dodatnio skorelowany z rozwojem unaczynienia tkanki nerwowej co również przemawia za mezodermalnym źródłem mikrogleju (Perry i in. 1985, Miyake i in. 1984, Hurley i in. 1996). Opuszczenie naczyń krwionośnych przez te komórki jest możliwe gdyż bariera krew-mózg tworzona w tym czasie przez astrocyty nie jest jeszcze dokładnie zamknięta w tym stadium życia organizmu. Po przeniknięciu dochodzi do transformacji mikrogleju amebowatego w mikroglej spoczynkowy charakteryzujący się obkurczoną częścią cytoplazmatyczna oraz licznymi rozgałęzieniami (Boya i in. 1991, Fedoroff i in. 1995).
Jako alternatywne źródło mikrogleju podaje się neuroektodermę, z której wywodzą się glioblasty będące prekursorami astrocytów i oligodendrocytów (Fujita i in. 1975,1980). Jednym z dowodów popierających teorię ektodermalnego pochodzenia mikrogleju jest fakt, że mikroglej, jak i komórki progenitorowe 0-2A, z których mogą się rozwinąć oligodendrocyty oraz astrocyty typu drugiego, posiadają jednakowe właściwości histochemiczne. Poza tym zidentyfikowano homologiczne epitopy powierzchniowe zlokalizowane na astrocytach, jak i na mikrogleju, a rozpoznawane przez przeciwciała monoklonalne LN-1 (Dickson i in. 1989). Zgodnie z neuroektodermalną teorią o pochodzeniu mikrogleju zakłada się, że mikroglej wykształcił się jako samodzielna linia z puli komórek macierzystych neurogleju. Faktem przemawiającym za tą teorią jest to, że glioblasty ze strefy okołokomorowej wykształconej z cewki nerwowej są prekursorami komórek Hortegi. Znaczyłoby to że oligodendrocyty, astrocyty i mikroglej są grupami siostrzanymi pochodzącymi z tej samej linii multipotencjalnych komórek neuroektodermalnych (Fujita i in. 1975,1980).
Ze względu na brak jednoznacznych i niepodważalnych faktów przemawiających za jedną z opcji pochodzenia mikrogleju proponowane jest alternatywne heterogeniczne jest źródło (De Groot i in.). Według tego założenia część mikrogleju zasiedlającego tkankę nerwową jest pochodzenia mezodermalnego, a część neuroektodermalnego. Bez względu na pochodzenie mikrogleju, przyjmuje on w mózgu formy i funkcje charakterystyczne tylko dla tego rodzaju komórek gleju w dużym stopniu zależne od stanu fizjologicznego tkanki.
Funkcja mikrogleju
Mikroglej stanowiący około 5-20% populacji komórek nieneuronalnych w mózgu występuje w warunkach normalnych w formie spoczynkowej. Główne funkcje fizjologiczne mikrogleju sprowadzają się do monitorowania mikrośrodowiska tkanki, usuwania umierających neuronów i reakcji w wyniku stwierdzenia obecności obcego antygenu (Nakajima i in. 1993). W formie nieaktywnej mikroglej posiada liczne i długie rozgałęzienia części cytoplazmatycznej komórki. Ta specyficzna struktura prawdopodobnie ułatwia potencjalną reakcję tych komórek jako specyficznego dla tkanki nerwowej elementu układu immunologicznego.
Charakterystyczna transformacja mikrogleju z formy spoczynkowej do aktywowanej została poznana i opisana przez Rio Hortege prawie wiek temu. W wyniku aktywacji dochodzi do przemian morfologicznych którym towarzyszy: zwiększenie rozmiaru ciała komórki oraz obkurczenie rozgałęzień. Na poziomie molekularnym dochodzi do ekspresji białek adhezyjnych, reorganizacji cytoszkieletu oraz ekspresja elementów kompleksu zgodności tkankowej MHC typu I jak i II (Perry i in. 1987, 1989, Aloisi i in. 2000).
Zaobserwowano, że aktywowanie mikrogleju może odgrywać rolę w ciężkich postaciach zaburzeń psychiatrycznych[2].
Mikroglej w uszkodzonej tkance nerwowej
W wyniku uszkodzenia tkanki nerwowej dochodzi w mózgu do odpowiedzi komórkowej i chemicznej otaczających komórek które to reakcje są efektem fizycznego naruszenia integralności tkanki nerwowej, jak i zmian w lokalnym mikrośrodowisku np. zmiany homeostazy jonowej. Jednocześnie z uszkodzeniem uruchamianych jest wiele procesów mających na celu doprowadzić z jednej strony do ochrony przed czynnikami uszkadzającymi z drugiej do naprawy powstałych zniszczeń tkanki (Berkenbosch i in. 1992).
Odpowiedzi komórkowej na czynnik uszkadzający w większości przypadków towarzyszy: proliferacja i migracja komórek mikrogleju i astrocytów, produkcja cytokin prozapalnych, funkcjonalne zmiany w śródbłonku naczyń krwionośnych oraz rekrutacja z krwiobiegu komórek układu immunologicznego w obręb uszkodzonej tkanki (Dong i in. 2001, Aloisi i in. 2001). W uszkodzonych neuronach dochodzi do ekspresji wczesnych genów odpowiedzialnych za stymulację i aktywowanie astrocytów i mikrogleju (Neumann i in. 2001, Raivich i in. 1999). A poprzez przerwaną ciągłość bariery krew-mózg aktywowane zostają monocyty z krwi obwodowej które na wzór mikrogleju ulegają następnie transformacji morfologicznej (Fujita i in. 1998, Leong i in. 1992, Maxwell i in. 1990).
Aktywacja mikrogleju jaką wywołuje uszkodzenie tkanki związana jest ze wzrostem poziomu czynników o funkcji immunologicznej. W okresie tym może dojść do indukcji ekspresji szerokiej rzeszy receptorów powierzchniowych które przyspieszają odpowiedź immunologiczną. Pośród nich są receptory biorące udział w rozpoznawaniu cząsteczek związanych z patogenem, receptory komplementu (np. CR1, CR3, CR4), receptory cytokin (np. TNFRI, TNFRII, IL-1RI, IL-12R) oraz chemokin (np. CCR2, CCR3, CXCR4, CX3CR1) jak i receptory ułatwiające interakcje z układem immunologicznym jak np. z limfocytami T czy immunoglobulinami (np. Fc RI, RII, RIII) (Aloisi i in. 2001, Perry i in. 1992, Nakajima i in. 1993).
W wyniku uszkodzenia CNS napływowe monocyty oraz mikroglej zaczynają produkować również wiele związków prozapalnych takich jak cytokiny (IL-1,TNFα, IL-6, IL-12, IL-15, IL –18), chemokiny (IL-8, fraktalkina, IP-10, MIP-1α, MIP-1β, MCP-1, RANTES, MDC) oraz związki cytotoksyczne (iNOS, wolne rodniki tlenowe i azotowe) i prostanoidy (PGD2, PGE2, tromboksan B2) Aloisi i in. 2001). Rezultatem tego działania jest wzmożony napływ elementów krwiopochodnych z krwi obwodowej takich jak kolejna fala monocytów i makrofagów oraz limfocytów T i B, czy neutrofile. W pierwszej fazie odpowiedzi immunologicznej inicjowanej przez takie cytokiny jak IL-1, IL-6 czy TNF dochodzi do synergicznej reakcji monocytów, makrofagów oraz mikrogleju. Fakt ten wskazuje na współzależność tych komórek w efekcie uszkodzenia tkanki nerwowej.
Zwiększona proliferacja, migracja a przede wszystkim silna fagocytoza to cechy mikrogleju oraz makrofagów sprzyjające naprawie uszkodzonych części tkanki. Wzmożona proliferacja mikroleju w obrębie uszkodzenia jest efektem interakcji z uszkodzonymi neuronami jak również wynikiem skomplikowanych oddziaływań ze strony astrocytów oraz autostymulacji mikrogleju (Aloisi i in. 2001, Neumann i in. 2001, Dong i in. 2001). W wyniku kontaktu z uszkodzonym neuronem lub degenerującym połączeniem neuronalnym mikroglej wykazuje zdolność do transformacji w fagocyta – makrofaga mającego za cel usunięcie pozostałości z miejsca uszkodzenia tkanki nerwowej.
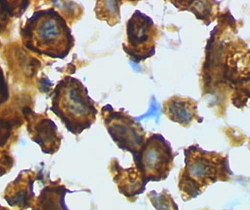
Właściwością aktywowanego mikrogleju służącą precyzyjnemu przeciwdziałaniu przyczynom oraz skutkom uszkodzenia mózgu jest zdolność tych komórek do transformacji w komórki prezentujące antygen (APC). Elementem kluczowym w pełnieniu tej funkcji przez komórkę mikrogleju jest zdolność do produkcji przez nią cząsteczki powierzchniowej CD40 oraz ekspresji elementów kompleksu zgodności tkankowej MHC typu I jak i II. Systemy oddziaływań CD40 – CD40L, MHC-II – TCR uważane są za kluczowe elementy w rozwoju odpowiedzi immunologicznej. Interakcje te mogą doprowadzić do rozwoju i różnicowania się aktywowanych limfocytów B czy też do pełnej aktywacji limfocytów T poprzez ekspresję cząsteczek stymulujących co ma miejsce w przypadku MS i EAE (Perry i in. 1987, 1989, Aloisi i in. 2000,2001).
W rezultacie odpowiedzi immunologicznej ze strony aktywowanego mikrogleju dochodzi z jednej strony do indukcji kaskady procesów naprawczych w obrębie uszkodzenie tkanki nerwowej. Z drugiej jednak strony procesowi naprawy towarzyszy wtórne uszkodzenie tkanki nerwowej a szczególnie neuronów co jest efektem nadmiernej produkcji cytokin prozapalnych (np. TNFα) produkowanych przez aktywowany mikroglej. Czynnikiem cytotoksycznym jest również produkcja wolnych rodników tlenowych jak i azotowych przez fagocyty usuwające pozostałości pierwotnego uszkodzenia.
Przypisy
- ↑ Kazuyuki Nakajima, Shinichi Kohsaka. Functional roles of microglia in the brain. „Neurosci Res.”. 17 (3), s. 187-203, August 1993. DOI: 10.1016/0168-0102(93)90047-T. [dostęp 2017-04-30]. (ang.).
- ↑ Mondelli, Valeria, Vernon, Anthony C, Turkheimer, Federico et al. Brain microglia in psychiatric disorders. „Lancet Psychiatry.”, 2017 Apr 25. DOI: 10.1016/S2215-0366(17)30101-3. [dostęp 2017-04-30]. (ang.).
Bibliografia
- K.K. Nakajima K.K., S.S. Kohsaka S.S., Functional roles of microglia in the brain, „Neuroscience Research”, 17 (3), 1993, s. 187–203, DOI: 10.1016/0168-0102(93)90047-T, ISSN 0168-0102, PMID: 8233123 .
- V.H.V.H. Perry V.H.V.H., D.A.D.A. Hume D.A.D.A., S.S. Gordon S.S., Immunohistochemical localization of macrophages and microglia in the adult and developing mouse brain, „Neuroscience”, 15 (2), 1985, s. 313–326, ISSN 0306-4522, PMID: 3895031 .
- TadahisaT. Kitamura TadahisaT., ToshihikoT. Miyake ToshihikoT., SetsuyaS. Fujita SetsuyaS., Genesis of resting microglia in the gray matter of mouse hippocampus, „The Journal of Comparative Neurology”, 226 (3), 1984, s. 421–433, DOI: 10.1002/cne.902260310, ISSN 1096-9861, PMID: 6747031 (ang.).
- S.D.S.D. Hurley S.D.S.D. i inni, Cytokine transcripts expressed by microglia in vitro are not expressed by ameboid microglia of the developing rat central nervous system, „Glia”, 25 (3), 1999, s. 304–309, ISSN 0894-1491, PMID: 9932876 [dostęp 2017-06-04] .
- JJ. Boya JJ. i inni, A lectin histochemistry study on the development of rat microglial cells., „Journal of Anatomy”, 175, 1991, s. 229–236, ISSN 0021-8782, PMID: 2050568, PMCID: PMC1224482 .
- S.S. Fedoroff S.S., C.C. Hao C.C., Origin of microglia and their regulation by astroglia, „Advances in Experimental Medicine and Biology”, 296, 1991, s. 135–142, ISSN 0065-2598, PMID: 1781324 .
- S.S. Fujita S.S., T.T. Kitamura T.T., Origin of brain macrophages and the nature of the so-called microglia, „Acta Neuropathologica. Supplementum”, Suppl 6, 1975, s. 291–296, ISSN 0065-1435, PMID: 168722 .
- S.S. Fujita S.S., Cytogenesis and pathology of neuroglia and microglia, „Pathology, Research and Practice”, 168 (4), 1980, s. 271–278, DOI: 10.1016/S0344-0338(80)80269-X, ISSN 0344-0338, PMID: 7413517 .
- D.W.D.W. Dickson D.W.D.W., L.A.L.A. Mattiace L.A.L.A., Astrocytes and microglia in human brain share an epitope recognized by a B-lymphocyte-specific monoclonal antibody (LN-1)., „The American Journal of Pathology”, 135 (1), 1989, s. 135–147, ISSN 0002-9440, PMID: 2476034, PMCID: PMC1880225 .
- Corline J. deC.J. Groot Corline J. deC.J., Christine D.Ch.D. Dijkstra Christine D.Ch.D., TaedeT. Sminia TaedeT., Discrimination between different types of neuroglial cells in rat central nervous system using combined immuno- and enzyme-histochemical methods, „Immunobiology”, 178 (3), 1988, s. 177–190, DOI: 10.1016/S0171-2985(88)80063-9, ISSN 0171-2985, PMID: 2465992 .
- B. HughB.H. Perry B. HughB.H., SiamonS. Gordon SiamonS., Modulation of CD4 antigen on macrophages and microglia in rat brain, „The Journal of Experimental Medicine”, 166 (4), 1987, s. 1138–1143, DOI: 10.1084/jem.166.4.1138, ISSN 0022-1007, PMID: 2443599, PMCID: PMC2188707 .
- V.H.V.H. Perry V.H.V.H., S.S. Gordon S.S., Macrophages and microglia in the nervous system, „Trends in Neurosciences”, 11 (6), 1988, s. 273–277, DOI: 10.1016/0166-2236(88)90110-5, ISSN 0166-2236, PMID: 2465626 .
- F.F. Aloisi F.F., The role of microglia and astrocytes in CNS immune surveillance and immunopathology, „Advances in Experimental Medicine and Biology”, 468, 1999, s. 123–133, ISSN 0065-2598, PMID: 10635024 .
- F.F. Berkenbosch F.F., Macrophages and astroglial interactions in repair to brain injury, „Annals of the New York Academy of Sciences”, 650, 1992, s. 186–190, DOI: 10.1111/j.1749-6632.1992.tb49119.x, ISSN 0077-8923, PMID: 1605474 .
- F.F. Aloisi F.F., Immune function of microglia, „Glia”, 36 (2), 2001, s. 165–179, ISSN 0894-1491, PMID: 11596125 .
- H.H. Neumann H.H., Control of glial immune function by neurons, „Glia”, 36 (2), 2001, s. 191–199, ISSN 0894-1491, PMID: 11596127 .
- GennadijG. Raivich GennadijG., Like cops on the beat: the active role of resting microglia, „Trends in Neurosciences”, 28 (11), 2005, s. 571–573, DOI: 10.1016/j.tins.2005.09.001, ISSN 0166-2236, PMID: 16165228 .
- T.T. Fujita T.T. i inni, Cellular dynamics of macrophages and microglial cells in reaction to stab wounds in rat cerebral cortex, „Acta Neurochirurgica”, 140 (3), 1998, s. 275–279, ISSN 0001-6268, PMID: 9638265 .
- Seng-KeeS.K. Leong Seng-KeeS.K., Eng-AngE.A. Ling Eng-AngE.A., Amoeboid and ramified microglia: their interrelationship and response to brain injury, „Glia”, 6 (1), 1992, s. 39–47, DOI: 10.1002/glia.440060106, ISSN 0894-1491, PMID: 1380949 .
- William L.W.L. Maxwell William L.W.L. i inni, Thalamic nuclei after human blunt head injury, „Journal of Neuropathology and Experimental Neurology”, 65 (5), 2006, s. 478–488, DOI: 10.1097/01.jnen.0000229241.28619.75, ISSN 0022-3069, PMID: 16772871 .
- V.H.V.H. Perry V.H.V.H., P.B.P.B. Andersson P.B.P.B., The inflammatory response in the CNS, „Neuropathology and Applied Neurobiology”, 18 (5), 1992, s. 454–459, DOI: 10.1111/j.1365-2990.1992.tb00811.x, ISSN 0305-1846, PMID: 1454134 .
- KazuyukiK. Nakajima KazuyukiK. i inni, Microglia-derived elastase produces a low-molecular-weight plasminogen that enhances neurite outgrowth in rat neocortical explant cultures, „Journal of Neurochemistry”, 61 (6), 1993, s. 2155–2163, DOI: 10.1111/j.1471-4159.1993.tb07454.x, ISSN 0022-3042, PMID: 8245967 .
| Zobacz multimedia związane z tematem: Mikroglej |
Kontrola autorytatywna (rodzaj komórki):
- Britannica: science/microglia
- DSDE: mikroglia












